Eredeti cikk dátuma: 2021. március 12.
Eredeti cikk címe: Persistence of SARS-CoV-2-specific B and T cell responses in convalescent COVID-19 patients 6–8 months after the infection
Eredeti cikk szerzői: Natalia Sherina (15), Antonio Piralla (15), Likun Du (15), Hui Wan (15), Makiko Kumagai-Braesch, Juni Andréll, Sten Braesch-Andersen, Irene Cassaniti, Elena Percivalle, Antonella Sarasini, Federica Bergami, Raffaella Di Martino, Marta Colaneri, Marco Vecchia, Margherita Sambo, Valentina Zuccaro, Raffaele Bruno, Michele Sachs, Tiberio Oggionni, Federica Meloni, Hassan Abolhassani, Federico Bertoglio, Maren Schubert, Miranda Byrne-Steele, Jian Han, Michael Hust Yintong Xue (16), Lennart Hammarström (16), Fausto Baldanti (16), Harold Marcotte (16), Qiang Pan-Hammarström (16, 17)
Eredeti cikk elérhetősége: https://www.cell.com/med/fulltext/S2666-6340(21)00038-6?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS2666634021000386%3Fshowall%3Dtrue
Eredeti cikk státusza: megjelent
Fordító(k): Czókolyová Monika
Lektor(ok): dr. Serly Julianna
Nyelvi lektor(ok): Csornainé Nagy Ágnes
Szerkesztő(k): dr.Szabó Edit, Vinkovits Mária
Figyelem! Az oldalon megjelenő cikkek esetenként politikai jellegű megnyilvánulásokat is tartalmazhatnak. Ezek nem tekinthetők a fordítócsoport politikai állásfoglalásának, kizárólag az eredeti cikk írójának véleményét tükrözik. Fordítócsoportunk szigorúan politikamentes, a cikkekben esetlegesen fellelhető politikai tartalommal kapcsolatosan semmiféle felelősséget nem vállal, diskurzust, vitát, bizonyítást vagy cáfolatot nem tesz közzé.
Az oldalon található információk nem helyettesítik a szakemberrel történő személyes konzultációt és kivizsgálást, ezért kérjük, minden esetben forduljon szakorvoshoz!
A tanulmányból kiemelendő
- A COVID19-betegminták többségénél SARS-CoV-2-elleni antitestek voltak jelen
- Az RBD- és S-specifikus IgG-szintek a diagnózis után 6 hónapig stabilak maradtak
- Specifikus memória B- és T-sejtek alakultak ki a COVID19-betegek több mint 95%-ánál
- A memória B- és T-sejt-válaszok a fertőzés után legalább 6–8 hónapig fennmaradtak
Kontextus és jelentőség
A 2019-es koronavírus-betegségből (COVID19) felgyógyult betegeknél az adaptív immunválasz hosszát vizsgáló tanulmányok hozzájárulhatnak ahhoz, hogy megértsük, hogyan alakul ki és marad fenn az immunvédelem a súlyos akut légzőszervi szindrómát okozó koronavírus 2 (SARS-CoV-2)-fertőzés természetes lefolyása alatt, így hasznos információt nyújtanak az új vírussal szembeni vakcinák értékeléséhez. Habár a SARS-CoV-2-immunglobulin G (IgG) antitestek szérumszintje szignifikánsan csökkent a fertőzést követően 6 hónappal, a gyógyult betegek többségében kialakultak a vírusspecifikus T- és/vagy memória B-sejt-válaszok, és viszonylag magasabb szinten fennmaradtak 6–8 hónappal a tünetek kezdete után. Ezek az adatok arra utalnak, hogy a természetes SARS-CoV-2-fertőzést követő védő adaptív immunitás legalább 6–8 hónapig fennmaradhat.
Összefoglalás
Háttér
Az adaptív immunválaszok monitorozása a súlyos akut légzőszervi tünetegyüttest okozó koronavírus (SARS-CoV-2)-fertőzés természetes lefolyása alatt hasznos információt nyújt a vírus és az újonnan felbukkanó variánsai elleni oltási stratégiák kidolgozásához. Ezért profiloztuk a szérum SARS-CoV-2-ellenes antitestszintet és a specifikus memória B- és T-sejt-válaszokat a 2019-es koronavírus-betegségből (COVID19) felgyógyult betegeknél.
Módszerek
Összesen 119 mintában vizsgáltuk a tüske és receptorkötő domén elleni megemelkedett antitestszintek jelenlétét egy 8 hónapos periódusban; a minták 88 donortól származtak, akik az enyhétől a kritikus lefolyásúig terjedő betegségből gyógyultak fel. Ezenkívül a SARS-CoV-2 neutralizáló antitestek szintjét és a specifikus memória B- és T-sejt-válaszokat teszteltük a minták egy alcsoportjában.
Eredmények
A SARS-CoV-2-ellenes antitestek a COVID19-betegek tüneteinek megjelenésétől számított 4 héten belül gyűjtött minták 85%-ában voltak jelen. A specifikus immunglobulin M (IgM)/IgA-antitestek szintje 1 hónap elteltével csökkent, míg a specifikus IgG antitestek szintje és plazmaneutralizáló aktivitása a diagnózis után 6 hónapig viszonylag stabil maradt. A SARS-CoV-2-ellenes IgG-antitestek továbbra is jelen voltak, habár szignifikánsan alacsonyabb szinten, a tünetek megjelenése után 6–8 hónappal gyűjtött minták 80%-ában. A SARS-CoV-2-specifikus memória B- és T-sejt-válaszok idővel kialakultak, és minden 6–8 hónapig követett betegnél állandóak voltak.
Következtetések
Adataink arra utalnak, hogy a természetes SARS-CoV-2-fertőzést követő védő adaptív immunitás a betegség súlyosságától függetlenül legalább 6–8 hónapig fennmaradhat. Így lehetségessé válhat a közép vagy hosszú távú védőimmunitás kialakítása védőoltással.
Pénzügyi támogatás
A projekt létrejöttét az Európai Unió Horizont 2020 kutatási és innovációs keretprogramja (ATAC, 101003650), az Italian Ministry of Health (olasz egészségügyi minisztérium, Ricerca Finalizzata, támogatás száma: GR-2013-02358399), a Center for Innovative Medicine (innovatív orvosi központ), és a Swedish Research Council (svéd kutatási tanács) támogatta. J.A. a SciLifeLab/KAW a 2020-as nemzeti COVID19 kutatási program projektjének támogatásában részesült.
Grafikus absztrakt

Kulcsszavak
- SARS-CoV-2
- COVID19
- immunitás
- antitest
- T-sejt
- B-sejt
- immunológiai memória
- az immunválasz hossza
- IgG
- neutralizáló antitest
Bevezetés
A súlyos akut légzőszervi tünetegyüttest okozó koronavírus 2 (SARS-CoV-2) megjelenése és terjedése pandémiához vezetett, amely jelentős hatással van az egészségre. Az új típusú koronavírus genomja 4 fő szerkezeti fehérjét kódol, ide értve a tüskefehérjét (S), a nukleoproteint (N), a membránfehérjét (M), a burokfehérjét (E), valamint más fehérjéket (ORF3a, ORF7a).1
Nagymértékű adaptív immunválaszt fedeztek fel a fertőzésből felépült betegekben, amelyben az S-specifikus neutralizáló antitestek, a memória B-sejtek és a keringő follikuláris segítő T-sejtek is részt vettek.2,3,4
Továbbá nagyszámú S-fehérje receptorkötő doménre (RBD) specifikus monoklonális antitestet izoláltak gyógyult betegektől származó mintákból, amelyeket állatmodellekben3,5,6 és klinikai vizsgálatokban elemeztek egy lehetséges passzív immunterápia kifejlesztése céljából.7,8,9
Ugyanakkor még mindig nem világos, meddig tart az adaptív immunitás a természetes SARS-CoV-2-fertőzés után, illetve, hogy az antitesttiter a védőimmunitás markerének tekinthető-e. Rézuszmakákóknál a SARS-CoV-2-fertőzés következtében kialakuló humorális immunválasz és az újra fertőződéssel szembeni védelem között összefüggést mutattak ki,10,azonban ezt az összefüggést emberben még nem határozták meg. Míg egy nemrégiben végzett izlandi vizsgálat azt mutatta, hogy az antitestválasz a gyógyult betegek 90%-ában a betegség kezdetét követően >4 hónappal is fennmaradt,11 más tanulmányok a betegség enyhe lefolyását mutató betegeknél a SARS-CoV-2-ellenes immunglobulin G (IgG) gyors lebomlását jelentették.12,13
Mindazonáltal a hosszú életű memória T- és B-sejtek jelen lehetnek és reaktiválódhatnak egy második expozíció alkalmával, ezáltal biztosítva az immunvédelmet.
A 2019-es koronavírus-betegségből (COVID19) felgyógyult betegeknél az adaptív immunválasz hosszát vizsgáló tanulmányok hozzájárulhatnak ahhoz, hogy megértsük, hogyan alakul ki és marad fenn az immunvédelem a SARS-CoV-2-fertőzés természetes lefolyása során, valamint hasznos információt nyújthatnak a vakcinák kifejlesztése és értékelése szempontjából az új vírussal szemben. Ebben a tanulmányban célunk a SARS-CoV-2-specifikus immunválaszok dinamikájának és hosszának értékelése olyan COVID19-betegek estében, akik a betegség széles spektrumán helyezkednek el. 88 betegtől, a kezdeti tüneteket követő 8 hónapban, különböző időpontokban vett mintákban értékeltük a SARS-CoV-2-specifikus antitestek szintjét és Ig-csoportját, valamint a memória B- és T-sejtek kialakulását.
Eredmények
A SARS-CoV-2-ellenes antitestválasz dinamikája COVID19-betegeknél
A SARS-CoV-2-fertőzés ellen kialakuló antitestválasz értékelése 88 COVID19-betegtől származó, összesen 119 szérum vagy plazmaminta (78 olaszországi és 10 svédországi beteg 1. ábra; 1. és S1 táblázat) vizsgálata saját fejlesztésű enzimhez kapcsolt immunoszorbens eljárással (ELISA) történt az anti-S és az anti-RBD antitest jelenlétének kimutatására (a részletékért lásd a Módszerek fejezetet). A SARS-CoV-2-specifikus IgM-, IgA- és IgG-antitestet COVID19-betegek 55 mintájában vizsgáltuk meg, amelyeket a gyógyulás korai szakaszában gyűjtöttünk (7–28 nappal a betegség tüneteinek megjelenése után), illetve 108 historikus kontrollban (a SARS-CoV-2-pandémia előtt gyűjtött minták). A betegeknél jelentősen magasabb anti-S és anti-RBD IgM- és IgA-antitestszinteket (minden csoportnál: P < 0,0001) mutattunk ki a historikus kontrollokhoz képest (2A–2C ábra). Az anti-S IgM-, IgA- és IgG-szintek a betegek 76, 78 és 85%-ában voltak emelkedettek, ebben a sorrendben. Az anti-RBD IgM-, IgA- és IgG-szintek a betegek hasonló arányában voltak emelkedettek, azaz 67%-ban, 62%-ban és 85%-ban voltak pozitívak. Az anti-S és anti-RBD antitesttiterek szorosan korrelálnak a különböző izotípusokkal (IgM: r = 0,88; IgA: r = 0,77; IgG: r = 0,95) (2D–2F ábra).

| Olasz kohorsz | Svéd kohorsz | |
| Demográfiai adatok | ||
| Szám | 78 | 10 |
| Életkor, év, medián (terjedelem) | 63,0 (32–89) | 53,5 (29–75) |
| Férfiak, % | 58 (45/78) | 50 (5/10) |
| Nők, % | 42 (33/78) | 50 (5/10) |
| A betegség súlyossága, % | ||
| Enyhe | 6 (5/78) | 100 (10/10) |
| Középsúlyos | 21 (16/78) | 0 (0/10) |
| Súlyos | 67 (52/78) | 0 (0/10) |
| Kritikus | 6 (5/78) | 0 (0/10) |
| Tünetek, % | ||
| Láz | 96 (75/78) | 40 (4/10) |
| Köhögés | 67 (52/78) | 40 (4/10) |
| Légszomj | 47 (37/78) | 0 (0/10) |
| Gyengeség | 12 (9/78) | 60 (6/10) |
| Hasmenés | 9 (7/78) | 0 (0/10) |
| Szaglásvesztés | 4 (3/78) | 50 (5/10) |
| Hipoxia | 1 (1/78) | 0 (0/10) |
| Orvosi anamnézis, % | ||
| Hipertónia | 51 (40/78) | 0 (0/10) |
| Cukorbetegség | 21 (16/78) | 0 (0/10) |
| Szívbetegség | 15 (12/78) | 0 (0/10) |
| Elhízás | 13 (10/78) | 0 (0/10) |
| HCV | 10 (8/78) | 0 (0/10) |
| Tüdőbetegségek | 5 (4/78) | 0 (0/10) |
| Daganatos megbetegedés | 5 (4/78) | 0 (0/10) |
| Egyéb társbetegségek | 41 (32/78) | 10 (1/10) |
| >1 társbetegség | 49 (38/78) | 0 (0/10) |
| >2 társbetegség | 29 (23/78) | 0 (0/10) |
| Súlyosság, % | ||
| Oxigénterápia | 73 (57/78) | 0 (0/10) |
| ITO | 6 (5/78) | 0 (0/10) |
1. táblázat | A COVID19-pozitív betegek demográfiai és klinikai jellemzőinek összegzése
HCV, hepatitisz C vírus; ITO, intenzív terápiás osztály. Lásd még az S1 táblázatot.
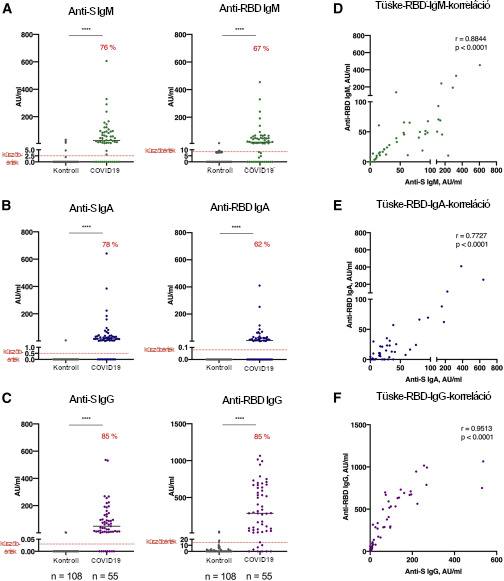
(A–C) Az anti-S és anti-RBD IgM-, IgA- és IgG-antitestszintek historikus kontrolloknál és COVID19-betegeknél.
(D–F) Az anti-S és anti-RBD IgM-, IgA- és IgG-titerek korrelációja.
Az antitesteket 55 COVID19-beteg mintájában mértük, amit a gyógyulás korai szakaszában gyűjtöttünk (7–28 nappal a betegség tüneteinek megjelenése után), illetve 108 historikus kontrollban (a SARS-CoV-2-pandémia előtt gyűjtött minták) mértük. A szimbólumok a vizsgálati alanyokat jelölik, a horizontális fekete vonalak pedig a mediánt mutatják. A szaggatott piros vonal jelzi az emelkedett anti-S és anti-RBD antitestszintek küszöbértéket (IgM esetén 2,5 és 8,4 AU/ml, IgA esetén 0,5 és 0,08 AU/ml, és IgG esetén 0,03 és 14,81 AU/ml), amelyeket a historikus kontrollok (n = 108) és a COVID19-betegek (n = 55) antitestválaszai alapján definiáltunk hatásfokmérő karakterisztika (ROC) görbe alkalmazásával.
A százalékos értékek (A–C) az antitestpozitívak gyakoriságát mutatják. Mann–Whitney U-próba (A–C) és Spearman-rangkorreláció (D–F). ∗∗∗∗P<0,0001.
A COVID19-diagnózis felállításakor fennálló tünetek alapján a betegeket enyhe, középsúlyos, súlyos vagy kritikus betegségstádiumba soroltuk (lásd részletesen: Módszer fejezet). A SARS-CoV-2-ellenes antitestszintek elemzése nem mutatott statisztikailag szignifikáns különbséget a súlyos és kritikus, illetve az enyhe és középsúlyos betegek csoportjainak összehasonlításakor (anti-S IgG esetén P = 0,1444 és P = 0,2943, anti-RBD IgG esetén P = 0,1203 és P = 0,4672) (3A és 3B ábra). Érdekes módon 6 betegben (11%) különösen alacsony anti-S és anti-RBD IgM-, IgA- és IgG-antitestszintek vagy ezen antitestek hiánya volt megfigyelhető. Az említett betegek szérummintáinak levétele a tünetek kezdete utáni 10,5. medián napon történt (terjedelem 7–22). A későbbi időpontban levett második minta 2 beteg esetében volt elérhető (86–226 nappal a tünetek kezdete után). Mindkét beteg anti-S- és anti-RBD IgG-pozitívvá vált. A maradék 4 beteg esetében (7%) nem állt rendelkezésre második minta az elemzéshez. Ennek a 4 betegnek magasabb volt a mediánéletkora (79 év) a teljes betegkohorszhoz viszonyítva (62 év), valamint betegségük súlyos besorolást kapott és később 2 beteg a COVID19 szövődményei miatt elhunyt. A betegek nem szerinti csoportosításakor egyedül az anti-RBD IgG esetében volt megfigyelhető statisztikailag szignifikáns különbség, ahol a férfiakban jelentősen magasabb IgG-szintek voltak jelen a súlyos/kritikus állapotú nőkkel összehasonlítva (P = 0,0306), az egyéb anti-S vagy anti-RBD-antitestszintek esetében nem volt megfigyelhető szignifikáns különbség (3C és 3D ábra).

Az anti-S és anti-RBD IgM-, IgA- és IgG-antitestszintek a betegség súlyossága (A és B) és a nemek szerint (C és D). A szimbólumok a vizsgálati alanyokat jelölik, a horizontális fekete vonalak pedig a mediánt mutatják. A szaggatott piros vonal az emelkedett anti-S és anti-RBD antitestszintek küszöbértékét jelzi (a 2. ábrához hasonlóan). Mann–Whitney U-próba. ∗P<0,05. Semmilyen statisztikailag szignifikáns különbség nem volt megfigyelhető (P>0,05) az anti-S (A) és anti-RBD (B) szintekben a súlyos és kritikus, illetve az enyhe (anti-S IgG esetén P = 0,1444 és P = 0,2943) és középsúlyos (anti-RBD IgG esetén P = 0,1203 és P = 0,4672) betegcsoportok összehasonlításakor.
Az SARS-CoV-2-ellenes antitestválasz hosszának vizsgálatakor tovább elemeztük a 88 betegtől különböző időpontokban (7–240 nappal a tünetek kezdete után) gyűjtött összesen 119 mintát. Az anti-S és anti-RBD antitestszintek már 7–14 nappal a tünetek kezdete után jelentősen megemelkedtek (P<0,0001 minden izotípusra), és a maximumot 15–28 nap között érték el (4. ábra) (IgM esetén P = 0,0087 és P = 0,0309, IgA esetén P = 0,0080 és P = 0,0029, IgG esetén P = 0,0159 és P = 0,0049, 7–14-től 15–28 napig). Az anti-S és anti-RBD IgM- és IgA-antitestszintek szignifikáns csökkenése volt megfigyelhető a 28. nap után (anti-RBD IgA esetén a 15–28. és a 29–90. napok között P = 0,0184; anti-S és anti-RBD IgM és IgA esetén a 15–28. és a 91–180. napok között P<0,0001, és a 15–28. és a 181–240. napok között P<0,0001). A 91–180. napra az anti-S és anti-RBD IgG szintekben szignifikáns csökkenés nem jelentkezett (a 15–28. és a 91–180. napok között anti-S IgG esetén P = 0,1847 és anti-RBD IgG esetén P = 0,0544), ugyanakkor a 181–240. napra szignifikáns csökkenés volt megfigyelhető (a 15–28. és a 181–240. napok között anti-S IgG esetén P = 0,0003 és anti-RBD IgG esetén P = 0,0002). Fontos megjegyezni, hogy a tünetek kezdete után 181–240 napig (6–8 hónap) követett betegek 80%-ában (12/15) jelentős SARS-CoV-2-elleni IgG-antitestválasz volt jelen. A diagnózis felállításakor ezek a betegek enyhe (n = 2), középsúlyos (n = 5) és súlyos (n = 5) lefolyású betegség jeleit mutatták.

4. ábra | SARS-CoV-2-ellenes antitestválasz hossza COVID19-betegeknél
Az anti-S és anti-RBD IgM-, IgA- és IgG-antitestválasz COVID19-betegekben a diagnózist és a felépülést követően (A és B). 88 betegtől összesen 119 mintát gyűjtöttünk. A mintákat 5 vizsgálati periódusban gyűjtöttük: 7–14. nap (n = 19), 15–28. nap (n = 36), 29-90. nap (n = 22), 91–180. nap (n = 27) és 181–240. nap (n = 15) a tünetek kezdete után. A szimbólumok a vizsgálati alanyokat jelölik, a horizontális fekete vonalak pedig a mediánt mutatják. A szaggatott piros vonal az emelkedett anti-S és anti-RBD antitestszintek küszöbértékét jelzi (a 2. ábrához hasonlóan). A csoportok közti szignifikáns különbségeket az oszlopok tetején jelöltük. Mann–Whitney U-próba. ∗P≤0,05, ∗∗P≤0,001, ∗∗∗P≤0,001, és ∗∗∗∗P<0,0001.
Lásd még az S1 ábrát.
A SARS-CoV-2-ellenes antitestválasz dinamikájának további értékelése érdekében összehasonlítottuk 27 beteg illesztett mintájának antitestszintjét. Az első minta (TP1) gyűjtése a tünetek kezdete utáni 21. medián napon (terjedelem 7–64), míg a második illesztett minta (TP2) gyűjtése a 126. medián napon (terjedelem 57–234) történt. Ez az analízis az anti-S IgM- és IgA-szintek (P<0,0001 és P = 0,0008), valamint az anti-RBD IgM- és IgA-szintek (P = 0,0002 és P = 0,0052) szignifikáns csökkenését mutatta az illesztett második mintákban (5. ábra). Azonban az anti-S és anti-RBD IgG szignifikáns csökkenése nem volt megfigyelhető (P = 0,1551).

Az anti-S (A–C) és anti-RBD (D–F) IgM-, IgA- és IgG-antitestszintek 27 COVID19-betegnél első, TP1 (medián 21. nap, terjedelem 7–64 nap) és második, TP2 (medián 126. nap, terjedelem 57–234 nap) időpontokban csoportos (bal oldali panel) és egyéni szinten bemutatva (jobb oldali panel). A szimbólumok a vizsgálati alanyokat jelölik, a horizontális fekete vonalak pedig a mediánt mutatják. A görbe alatti szürke terület (jobb oldali panelek) az antitest bomlási rátáinak eloszlását mutatja, amelyet kétfázisú exponenciális asszociációval becsültünk meg. Wilcoxon-féle előjeles rangpróba. ∗∗P≤0,001 és ∗∗∗P≤0,001.
Végezetül a SARS-CoV-2-neutralizáló antitestszinteket mikroneutralizációs tesztekkel mértük 96 mintában (a részletekért lásd a Módszerek fejezetet). A neutralizáló antitesttiterek korreláltak az anti-S IgM (r = 0,42), anti-S IgA (r = 0,36) és anti-S IgG (r = 0,46) szintekkel, ahogyan az anti-RBD IgM (r =0,38), anti-RBD IgA (r = 0,21) és anti-RBD IgG (r = 0,42) szintekkel is (S1A és S1B ábra). Az anti-S és anti-RBD antitestek dinamikájához hasonlóan a plazma neutralizáló aktivitásai a maximumot a 15–28. napon érték el (S1C és S1D ábra). Továbbá az IgG-izotípushoz hasonlóan a neutralizáló antitesttiterek a fertőzés után stabilak maradtak 91–180 napig, és, bár jelentősen csökkentek, jelen voltak a 181–240. napon (S1C és S1D ábra) is. Ezek az adatok arra utalnak, hogy a vizsgálati kohorszunkban a COVID19-betegek többségében kiváltódott a SARS-CoV-2-ellenes antitestválasz, és habár az IgM- és IgA-antitestek gyorsan csökkentek, az IgG-antitestek és a plazmaneutralizáló aktivitás noha alacsonyabb szinten, de továbbra is jelen volt a diagnózist követően legalább 6–8 hónapig.
A SARS-CoV-2-specifikus memória B- és T-sejtek indukciója
Annak érdekében, hogy megválaszoljuk a kérdést, keletkeztek-e SARS-CoV-2-specifikus memória B- és T-sejtek és milyen hosszan volt jelen a B- és T-sejt közvetítette immunválasz a COVID19-betegekben, 32 perifériás vér mononukleáris sejt (PBMC) mintát elemeztünk, amelyek 24 betegtől származtak (enyhe = 11, középsúlyos = 4, súlyos = 8, kritikus = 1; S1 táblázat). 17 beteg esetén csupán egy időpontban, míg 6 betegnél 2 időpontban és 1 betegnél 3 időpontban történt mintavétel. A mintákat az időpontok alapján további 3 csoportra osztottuk (a tünetek kezdete után 14–35 nappal, 85–180 nappal és 181–236 nappal történt a mintavétel, ∼azaz 2–4 hét, 3–6 hónap és 6–8 hónap). Az ELISpot eljárással semennyi vagy elhanyagolható mennyiségű RBD-specifikus IgA-antitestet szekretáló B-sejtet detektáltunk (részletekért lásd a Módszerek fejezetet) a 4 egészséges kontrolltól származó mintában és a 7 pandémia előtti minta felülúszójában. A negatív kontrollok közül a legmagasabb megfigyelt értéket használtuk küszöbértékként. RBD-specifikus IgG-t termelő B-sejteket detektáltunk 2–4 héttel a tünetek kezdete után a betegektől gyűjtött minták 33%-ában (2/6), 3–6 hónappal a tünetek kezdete után gyűjtött minták 93%-ában (14/15), és 6–8 hónappal a tünetek kezdete után gyűjtött minták 100%-ában (11/11) (6A ábra). A korai időpontból származó mintákhoz hasonlítva az RBD-specifikus IgG-t termelő sejtek száma jelentősen magasabb volt a kései időpontokban (3–6 hónap P = 0,0001 és 6–8 hónap P = 0,0006 a 2–4 hónaphoz viszonyítva) (S2A ábra; S2 táblázat). Úgy tűnik, hogy a B-sejt-válasz a maximumát a fertőzést követő 3–6 hónappal érte el, habár szignifikáns csökkenés nem volt megfigyelhető 6–8. hónapban (6B és S2A ábra, P = 0,1982; S2 táblázat). Továbbá a specifikus B-sejtek számának vonatkozásában nem volt megfigyelhető szignifikáns különbség azon betegek között, akiknek a betegsége eltérő súlyosságú volt (S3A ábra; S3 táblázat). Így, bár az idő múlásával az anti-RBD IgG-szintek csökkenése volt megfigyelhető, a SARS-CoV-2-specifikus memória B-sejtek szinte minden, a vizsgálat végéig követett betegnél megjelentek és megmaradtak a betegség súlyosságától függetlenül (6A, 6B, S2A, S2B, S3A és S3B ábra).

(A) RBD-specifikus memória B-sejtek száma kontroll (n = 11) és COVID19-betegek (n = 32) mintáiban.
(B és F) RBD-specifikus memória B-sejtek, SARS-CoV-2-specifikus T-sejtek és szérum anti-RBD IgG-szintek dinamikája COVID19-betegek mintáiban az idő múlásával a lognormális görbeillesztésnek megfelelően.
(C–E) Az S1-re, az S2N-re, és az S N M O SARS-CoV-2-fehéjeeredetű peptidkészletekre specifikus, IL-2-t (C), IFN-γ-t (D), vagy IFN-γ-t és IL-2-t (E) termelő T-sejtek száma a kontrollokban és a COVID19-betegek mintáiban.
(G) Specifikus memória B-sejtek és az S1-re, S2N-re és az S N M O fehérjeeredetű peptidkészletekre specifikus IFN-γ-t/IL-2-t termelő T-sejtek száma 7 beteg mintájában (enyhe [CP45], középsúlyos [CP08, CP09], és súlyos [CP34, CP03, CP19, CP06] lefolyású betegség esetén) az 1., a 2. és a 3. (CP06-ra) időpontokban.
Az eredményeket a negatív kontroll hátterének levonása után 300 000 szélesztett sejtre jutó pöttyök számaként fejeztük ki. A piros vonal a csoport medián értékét jelzi. A küszöbértéket a negatív kontrolloknál a specifikus B- és T-sejtek legmagasabb mennyiségére állítottuk be. Mann–Whitney U-próba. ∗∗P≤0,01, ∗∗∗P≤0,001, és ∗∗∗∗P≤0,0001.
Lásd még az S2–S4 ábrát és az S2 és S3 táblázatot.
Továbbá, míg elhanyagolható mennyiségű vagy semennyi interleukin 2-t (IL-2), interferon-γ-t (IFN-γ) vagy IL-2-t/IFN-γ-t termelő T-sejtet nem lehetett kimutatni a három SARS-CoV-2 eredetű peptidkészlet ellen a FluoroSpot eljárással (részletekért lásd a Módszerek fejezetet) a negatív kontrollokban, a vizsgált betegminták 17–100%-ában figyeltünk meg a küszöbérték feletti számban ilyen T-sejteket a tesztelt peptidkészlettől függően (S1, S2 N, vagy S N M O fehérje eredetű) és ezekben vizsgáltuk a citokineket (6C–6E ábra). Két betegnél nagyszámú IFN-γ-t és/vagy IL-2-t termelő T-sejtet mutattunk ki a kontrolltenyészetben is a SARS-CoV-2-peptidek hozzáadása nélkül, ami folyamatban levő gyulladásra utalhat. Összességében a T-sejt-válasz legalább az egyik SARS-CoV-2-peptidkészlettel szemben kimutatható volt az összes korai időpontban (2–4 hét) tesztelt betegnél (n = 6), és a válasz a vizsgált betegek többségében fennmaradt a későbbi időpontokban is (93% [14/15] a tünetek kezdete után 3–6 hónappal és 100% [11/11] a tünetek kezdete után 6–8 hónappal). Érdekes módon az egyetlen betegnél, akinek nem volt T-sejt-válasza a 4. hónapban, kimutatható volt a B-sejt-válasz. A korai időpontból származó mintákhoz hasonlítva az S1-specifikus IL-2-t, IFN-γ-t, és IL-2-t/IFN-γ-t termelő T-sejtek száma jelentősen magasabb volt a későbbi időpontokból származó mintákban, különösen a fertőzés utáni 6–8. hónapban gyűjtött mintákban (P = 0,0047, P = 0,0033 és P = 0,0019; S2C–S2E ábra; S2 táblázat). Hasonló eredményeket lehetett megfigyelni az S N M O peptidkészlet-specifikus T-sejtek esetében is, de az S2-peptidkészlet-specifikus T-sejtek esetében nem. A három peptidkészlet esetében a citokint termelő T-sejtek számának vonatkozásában nem volt megfigyelhető statisztikailag szignifikáns különbség az eltérő súlyosságú betegek között (P>0,05; 3C–3E ábra; S3 táblázat). Míg az anti-RBD IgG-szintek idővel csökkentek, a SARS-CoV-2-specifikus T-sejtek a specifikus memória B-sejtek mellett szintén kialakultak és fennmaradtak mind a mai napig követett betegek többségében a betegség súlyosságától függetlenül (6F ábra).
A vírusspecifikus B- és T-sejt-válaszok dinamikájának további vizsgálata érdekében külön elemeztük a betegek legalább 2 időpontból származó mintáit (6G ábra). 7 betegből 6-ban a vírusspecifikus B- és T-sejtek számának emelkedése volt megfigyelhető a második időpontban (TP1 = 14–116 nap, TP2 = 124–236 nap; 6G ábra). A fennmaradó egy betegben (gyógyult beteg [CP]19) a vírusspecifikus B- és T-sejtek számának enyhe csökkenése volt megfigyelhető a 2. időpontban (TP1 = 122 nap, TP2 = 228 nap).
Végezetül, a SARS-CoV-2-specifikus adaptív immunitás szempontjából az összes eljárással (vírusspecifikus antitestek, memória B- és T-sejtek) értékelt betegnél ábrázoltuk az anti-RBD-specifikus IgM-, IgA- és IgG-szintek, az RBD-specifikus IgG-t termelő B-sejtek, és az S1-peptidkészlet-specifikus IL-2-t és/vagy IFN-γ-t termelő T-sejtek számának normalizált log2 értékeit minden résztvevő esetében (S4 ábra). Ugyanazon személyek különböző időpontokból származó mintáinak ábrázolása színkódolva történt (S4 ábra). Habár a legtöbb minta egyedi mintázatot mutat, a 24 betegből 6 és 17 betegnél legalább 1 időpontban az adaptív immunitás 2 vagy minden „karja” kialakult különböző szinteken, a fennmaradó egyetlen betegnél pedig csupán memória B-sejteket detektáltunk (CP10, mintavétel a 126. napon). Azon betegeknél, akiknek több mint 1 időpontból származik mintája, egyértelmű eltolódást figyeltünk meg a specifikus antitestek (beleértve az IgM-et/IgA-t a korai időpontban) termelésétől a memória B és T-sejtek keletkezése irányába a későbbi időpontokban (S4 ábra). Habár léteznek egyéni különbségek, a SARS-CoV-2-specifikus memória B- és T-sejtek kialakultak és fennmaradtak a vizsgált COVID19-betegek többségében, betegségük kezdeti súlyosságától függetlenül, ami arra utal, hogy a SARS-CoV-2 elleni adaptív immunitás természetes fertőzés esetén legalább 6–8 hónapig fennmarad.
Megbeszélés
A jelen tanulmányban az anti-S és anti-RBD IgM-, IgA- és IgG-antitestszinteket normalizálással, sorozathígításos pozitív referencia szérumkészlethez viszonyítva, valamint historikus kontrollminták alapján beállított küszöbérték mellett mértük meg. Adataink azt mutatták, hogy a SARS-CoV-2-ellenes antitestválasz a COVID19-betegek többségében már a tünetek kezdete után 2 héttel jelen volt, és az anti-S és anti-RDB IgG-szintje viszonylag stabil maradt a diagnózis után 6 hónappal, majd a 6–8. hónapban szignifikáns csökkenés következett be, míg az anti-S és anti-RDB IgM- és IgA-szintjének csökkenése már a betegség kezdete után 1–3 hónappal megfigyelhető volt. Az eredményeink összhangban vannak a korábbi vizsgálatokkal, amelyek hasonlóan hosszú élettartamot és SARS-CoV-2-ellenes antitestválasz-mintázatot mutatnak, ahol az antitestszintek a tünetek kezdete utáni 23. napon érik el a csúcsot és legalább 4 hónapig maradnak fenn,11, 14, 15, 16, 17, 18, 19, 20 más vizsgálatokkal viszont ellentmondásban vannak. Ezen vizsgálatokban az SARS-CoV-2-ellenes antitestek alacsony prevalenciája és gyors (3 hónapon belüli) eltűnése volt megfigyelhető enyhe vagy súlyos betegségben szenvedő COVID19-betegeknél.12,21 Más jelentésekkel egyetértésben az anti-RBD IgG-antitest magasabb titereit figyeltük meg azoknál a férfiaknál, akiket súlyosabban érintett a SARS-CoV-2-fertőzés, míg a vizsgálatunkban a súlyos betegségben szenvedő betegek 7%-ában nem alakult ki vagy extrém alacsony szinten maradt az antitestszint, ami arra utal, hogy gyengébb antivirális immunválasz alakult ki náluk.11,13,21 A specifikus antitestek magasabb szintje, vagy a spektrum másik vége (azaz a specifikus antitestek hiánya) súlyos betegségben szenvedők és férfi betegek esetében a magasabb vírusterhelés, a vírusürítés hosszabb időtartama22, 23 vagy más gazda/genetikai tényezőknek köszönhető.24, 25
A SARS-CoV-2-ellenes antitestválasz prevalenciáját és élettartamának hosszát illető korábban közölt ellentmondásos eredmények a SARS-CoV-2-elleni antitestek mérésére alkalmas standard eljárás hiányából következhetnek, mivel a közölt tanulmányok többsége különböző SARS-CoV-2-atigének (RBD-, S- és N-fehérjék) használatán alapul. A vizsgálatok eredményei közötti eltérések a COVID19-kohorszok közötti különbségekkel is magyarázhatók, mivel a betegek számánál figyelembe vették a mintagyűjtési időpontokat és az alkalmazott kezelést, valamint a vizsgálati alanyok életkorát és nemét, amelyek egyaránt befolyásolhatták az elemzést.26 Ezenkívül a negatív kontrollok használata és a küszöbértékek meghatározása is hozzájárulhat a vizsgálatok közötti különbségekhez. Vizsgálatunk során mind az enyhe, mind a súlyos betegségben szenvedő betegeket teszteltük RBD- és S-antigének felhasználásával, majd a betegeket historikus kontrollokhoz (prepandémiás minták) hasonlítottuk, és ennek megfelelően állítottuk be a küszöbértékeket. Ilyen körülmények között bizonyítottuk, hogy az anti-S és anti-RBD IgG-szintje viszonylag stabil maradt, azaz a diagnózis után 6 hónappal, a betegség súlyosságától függetlenül, nem történt szignifikáns csökkenés. Bár az antitestek védő szerepe a SARS-CoV-2-vel szemben továbbra is bizonytalan, a korábbi jelentésekkel összhangban megfigyeltük, hogy az anti-RBD-antitestszintek korrelálnak a plazma neutralizációs aktivitásával.17,19,27 Egy nemrégiben megjelent tanulmány szerint annak ellenére, hogy az anti-RBD, S2- és N-fehérje-antitestszintje a 6–8 hónapos időszak alatt csökkent, a neutralizációs titerek nem csökkentek szignifikáns mértékben.28 Vizsgálatunkban az anti-RBD és az anti-S IgG-antitestek 6 hónap elteltével megfigyelt csökkenése párhuzamosítva volt a megfelelő neutralizációs titerek csökkenésével, ami összhangban áll egy másik, nemrégiben készült jelentéssel.32 Fontos kiemelni, hogy az anti-RBD antitestek és a plazma neutralizációs aktivitása mindkét esetben csökkent, de a fertőzés után 6 hónappal is kimutatható maradt. Míg vizsgálati kohorszunk viszonylag kicsi, a különböző spektrumú betegséggel rendelkező olasz és svéd betegek bevonása és a hosszú utánkövetési idő (akár 6–8 hónap) segíthet megoldani a SARS-CoV-2-specifikus antitestek perzisztenciájáról folyó jelenlegi vitát. A két másik koronavírus, a SARS-CoV és a közel-keleti légzőszervi tünetegyüttes (MERS)-CoV elleni antitestek a fertőzés megjelenése után még 1–3 évvel is kimutathatóak voltak,22 ami arra utal, hogy a SARS-CoV-2-specifikus antitestek hosszabb ideig jelen lehetnek, mint ahogyan azt ezidáig megfigyeltük.
Az eddigi tanulmányok főként a specifikus antivirális antitestválasz élettartamának hosszára fókuszáltak. A B- és T-memóriasejtek kialakulása azonban kritikus a hosszú távú védelem szempontjából, és e memóriasejtek hosszú időtávú dinamikája továbbra is kevéssé ismert. Eredményeink azt mutatják, hogy a betegek többsége a betegség súlyosságától függetlenül képes olyan specifikus memória B-sejt-válaszokat kialakítani, amelyek a tünetek kezdete után legalább 6–8 hónapig fennmaradnak. Ezek az eredmények összhangban vannak a nemrégiben megjelent, főként enyhén vagy középsúlyosan érintett betegkohorszokon alapuló vizsgálatokkal (beleértve az elbírálás előtti közleményeket is), ahol az RBD-specifikus memória B-sejtek mérésére áramlási citometriás eljárásokat alkalmaztak.18,20,29,30,31,32 Kimutattuk azt is, hogy ezek a memória B-sejtek stimulálás hatására RBD-specifikus IgG-antitestek termelésére képesek.
Korábban kimutatták, hogy az S1- és más SARS-CoV-2-fehérjéből származó peptidek specifikus T-sejt-válaszokat indukálnak az enyhétől a súlyos lefolyású betegségben szenvedő betegeknél 1–3 hónappal a tünetek kezdete után.2,29,33,34,35 Az utóbbi időben arról is beszámoltak (beleértve az elbírálás előtti közleményeket is), hogy az elsődleges fertőzés után a SARS-CoV-2-specifikus T-sejtek legalább 6 hónapig fennmaradnak minden tesztelt COVID19-betegben, elsősorban az enyhe vagy középsúlyos betegkohorszokban.20,36,37,38 Eredményeink megerősítik és kiterjesztik a korábbi eredményeket, amelyek azt mutatják, hogy a SARS-CoV-2-specifikus IL-2-t és/vagy IFN-γ-t termelő sejtek jelen vannak az enyhétől a súlyos lefolyású betegségben szenvedő betegektől származó minták túlnyomó többségében. Továbbá arra jutottunk, hogy a specifikus T-sejt-válaszok nemcsak kimutathatók voltak, hanem a betegség megjelenése után 6–8 hónappal is fennmaradtak (a szakirodalomban fellelhető leghosszabb követési idő). Ez némileg ellentétben áll az utóbbi időben megjelent tanulmányokkal, amelyek a T-sejt-válasz csökkenését mutatják 6–8 hónappal a fertőzés után tünetmentes, és tüneteket mutató COVID19-betegeknél egyaránt.20,38 A különbséget a vizsgálatunkba bevont minták kis számával lehet magyarázni, különösen a betegség korai szakaszában, és/vagy a kohorszunkban lévő idősebb és súlyos betegek nagyobb arányával, mivel a SARS-CoV-2-specifikus T-sejt-válasz erősebb lehet és hosszabb ideig fennmaradhat súlyos lefolyású betegségben szenvedő betegeknél a magasabb vírusterhelés és/vagy a vírusantigének hosszabb perzisztenciája miatt.20,38 Egyes tanulmányokban azt is megfigyelték, hogy a SARS-CoV-2 S- és/vagy M-fehérje ellen irányuló T-sejt-válaszok az expozíció nélküli egészséges véradók 25–50%-ában jelen vannak, ez összhangban áll a nagyfokú, potenciálisan keresztreaktív T-sejt-immunitással az átlagpopulációban.2,34,35 A betegségben nem érintett kontrollokban nem észleltünk jelentős mennyiségű vírusellenes keresztreaktív T-sejtet, ennek okai az eltérő kísérleti körülmények, ideértve az alkalmazott negatív kontrollokat és egyes vizsgálatokban a hosszabb in vitro peptidekkel történő stimulációt39; azonban keresztreaktív T-sejteket a rövidebb ideig tartó in vitro stimulációt alkalmazó vizsgálatokban is megfigyeltek (9–24 óra).2,34,35
Az IL-2 és IFN-y termelésével és ezen citokinek kettős termelésével jellemzett S1-specifiikus T-sejtek kimutatása az effektor és proliferációs potenciállal rendelkező T-sejtek in vivo indukcióját jelzi. Az IFN-γ-t termelő T-sejtek az intracelluláris kórokozókkal szembeni immunitás legfőbb jellemzői, és bár tanulmányunkban nem vizsgáltuk a SARS-CoV-2-specifikus IFN-γ-t termelő T-sejteket, korábban kimutatták, hogy azok CD4+ (T-helper 1 [Th1]-szerű) vagy CD8+ citotoxikus fenotípusúak.2,34 Kimutatták, hogy azoknál a gyógyult betegeknél akiknél a SARS-CoV-2 S1-fehérjéjével szemben nem volt kimutatható antitest, a T-sejt-válaszaik erősebben irányultak az M-fehérje ellen, mint az S1-fehérje ellen.40 Ezenkívül az S-specifikus IFN-γ+ CD4+ T-sejtek Th1-alapú sejtes immunválaszát poolozott S-peptidek ellen mutatták ki az S-fehérjével oltott majmok többségében, és ez specifikus és neutralizációs anti-S-antitestek indukciójával társult.41 Eredményeink arra utalnak, hogy a vakcinához az S-fehérje immunogénként történő használata potenciálisan képes az emberekben az S-fehérje és az RBD-specifikus memória T- és B-sejtek indukálására.
Fontos, hogy bár vizsgálatunkban a szérum IgG-antitestszintek az idő múlásával csökkentek, a specifikus memória B- és T-sejtek a 6–8 hónapos időszakban fennmaradtak, és hozzájárulhattak a SARS-CoV-2-újra fertőződéssel szembeni védelemhez.29,31 Azonban a SARS-CoV-2-antitestek detektálása, ideértve az antitestek neutralizációját, valamint a memória B-sejtek és a T-sejtek hosszú ideig történő detektálását, nem feltétlenül jelzi a védő és a hosszú távú immunitást, és a védelemmel való összefüggése továbbra is meghatározásra vár. A közönséges humán koronavírusokról szóló vizsgálatok azt mutatják, hogy a neutralizációs antitestek indukálódnak, és egy nemrégiben közölt jelentés szerint a közönséges náthát okozó koronavírus elleni védő immunitás 6–12 hónapig tarthat.42 A szezonális koronavírusokkal történő újra fertőzés általában 3 éven belül történik,22 és az ismétlődő fertőzések általában enyhébb tünetekkel és kisebb vírusterheléssel társulnak.22,43 A tüskefehérjével szembeni neutralizációs antitestek egyszeri intravénás beadása enyhe vagy középsúlyos COVID19-betegeknél csökkenti a vírusterhelést és lerövidíti a hospitalizáció hosszát.8 Továbbá a SARS-CoV-2-fertőzés és az S-fehérje elleni védőoltás megvédheti a rézuszmakákókat a challenge (szándékos) fertőzéstől.10 Ezért valószínű, hogy az antitestek és a sejtes immunitás csökkenteni fogja a fertőzés kockázatát és csillapítja újra fertőződés esetén a betegség súlyosságát. Fentiek miatt tervezzük, hogy kibővítjük kohorszunkat és hosszabb ideig követjük nyomon, hogy értékelhessük az immunológiai memória fennmaradását.
Összegzésként elmondható, hogy a betegek többségében a SARS-CoV-2-specifikus memória B- és T-sejtek magas szintje 6–8 hónappal a fertőzés után arra utal, hogy a természetes fertőzés utáni immunitás legalább átmenetileg protektív lehet, és ezáltal lehetséges a közepes vagy hosszú távú védőimmunitás vakcináció általi kialakítása. Az S-fehérje epitópjaival szembeni T-sejt-reaktivitás és az RBD-doménnel szembeni antitestek felfedezése arra utal, hogy az S-fehérje felhasználásával az RBD-t célzó antitestek indukciójára szolgáló vakcina kifejlesztése megalapozott megközelítés.33 Jelenleg azonban nem világos, hogy a korábbi fertőzési hullámok során kialakult neutralizációs antitestek, és B- és T-sejt-válaszok valamint a jelenlegi oltási stratégiák védenek-e az új SARS-CoV-2-variánsokkal, például az 501Y.V2-vel (B.1.351, brit variáns) szemben.44 A SARS-CoV-2-vakcina kifejlesztése és értékelése iránti sürgős igény kielégítése érdekében javasoljuk, hogy a specifikus antitestválaszok és azok élettartama hosszának elemzése mellett fontolják meg a specifikus memória B- és T-sejtek mérését, amelyek a hosszú távú immunitás fő összetevői, valamint az újra fertőzés (azonos vagy eltérő variánsokkal) elleni védelemmel való összefüggés vizsgálatát.
A vizsgálat korlátai
Vizsgálatunknak vannak bizonyos korlátai, beleértve a beteg kohorsz kis méretét (119 minta 88 betegtől) és a T- és B-sejtek szempontjából elemzett minták kis számát (32 minta, beleértve a 7 főtől származó illesztett mintákat). Ezenkívül az első időpontból származó minták főként a lábadozás korai szakaszából származtak (a tünetek kezdete után 14–28 nappal), és kevesebb minta elemzése történt B- és T-sejtekre vonatkozóan a diagnózis után 1–3 hónappal. A korai időpontban mért specifikus T-sejtek szintje alacsonyabb volt, mint korábban a tünetek kezdete után 1–2 hónappal a gyógyult betegeknél mért érték, ami arra utal, hogy a memóriasejtek száma ebben az időszakban növekedhet.34,45 Továbbá a kísérleti körülmények miatt nem lehetett megkülönböztetni, hogy a korai időpontokban mért T-sejtek effektor- vagy memóriasejtek voltak-e. A memória B- és T-sejtek kialakulásának és perzisztenciájának értékeléséhez nagyobb minta értékelésére van szükség, amit több időpontban követnek a vizsgálati időszak alatt, valamint el kell végezni a sejtek fenotipizálását.
STAR Módszerek
| REAGENS vagy ERŐFORRÁS | FORRÁS | AZONOSÍTÓ |
| Antitestek | ||
| (HRP)-konjugált kecske anti-humán IgM | Invitrogen | Cat# A18835; RRID: AB_2535612 |
| (HRP)-konjugált kecske anti-humán IgA | Jackson | Cat# 109-036-011; RRID: AB_2337592 |
| (HRP)-konjugált kecske anti-humán IgG | Invitrogen | Cat# A18805; RRID: AB_2535582 |
| Baktérium- és vírustörzsek | ||
| SARS-CoV-2 | Betegizolátum (Olaszország) | N/A |
| Biológiai minták | ||
| Vérminták (konvaleszcens) | Olaszország, ez a cikk | S1 táblázat |
| Vérminták (konvaleszcens, egészséges) | Svédország, ez a cikk | S1 táblázat |
| Szérum (konvaleszcens, egészséges) | Svédország, ez a cikk | S1 táblázat |
| Felülúszó (egészséges donorok) | Svédország, ez a cikk | S1 táblázat |
| Vegyi anyagok, peptidek és rekombináns fehérjék | ||
| Ni-NTA gyanta | ThermoFisher | Cat# 88221 |
| 16/600 Superdex 200 kDa | Cytiva | Cat# 28989335 |
| Lymphoprep | Axis-Shield | Cat# 1114547 |
| RNeasy mini kit | QIAGEN | Cat# 74106 |
| imidazoquinolin rezikvimod | Mabtech AB | Cat# R848 |
| Tetrametilbenzidin szubsztrát | Sigma | Cat# T0440 |
| Tween 20 | Sigma | Cat# P9416 |
| PBS | Karolinska University Hospital | Cat# MIK3125-1000 |
| BSA | Sigma | Cat# A7906 |
| RPMI1640 | ThermoFisher Scientific | Cat# 61870010 |
| FBS | ThermoFisher Scientific | Cat# 10270106 |
| Penicillin-sztreptomicin | ThermoFisher Scientific | Cat# 15140122 |
| SARS-CoV-2 S1-peptidkészlet | Mabtech AB | Cat# 3629-1 |
| SARS-CoV-2 SNMO-peptidkészlet | Mabtech AB | Cat# 3622-1 |
| SARS-CoV-2 S2N-peptidkészlet | Mabtech AB | Cat# 3620-1 |
| RBD-His rekombináns fehérje | Saját fejlesztés, ez a cikk | N/A |
| S1-S2-His rekombináns fehérje | Saját fejlesztés, ez a cikk | N/A |
| Kritikus fontosságú kereskedelemben elérhető tesztek | ||
| RBD ELISpot PLUS (ALP) kit | Mabtech AB | Cat# 3850-4APW-R1-1 |
| Human IFN-γ/IL-2 FluoroSpot PLUS kit | Mabtech AB | Cat# FSP-0102-2 |
| Kísérleti modellek: sejtvonalak | ||
| Expi293 | ThermoFisher Scientific | Cat# A14527; RRID: CVCL_D615 |
| High Five rovarsejtek BTI-TN-5B1-4 | ThermoFisher Scientific | Cat# B85502; RRID: CVCL_C190 |
| Vero-E6 | ATCC | Cat# CRL-1586; RRID: CVCL_0574 |
| Szoftver és algoritmusok | ||
| GraphPad Prism 7 és 8 | GraphPad szoftver | https://www.graphpad.com/ |
| R 3.6.1-es verzió | RStudio | https://rstudio.com/ |
| Egyéb | ||
| High-binding half area lapos aljú mikrotiter lemezek | Corning | Cat# 3690 |
Erőforrások elérhetősége
Vezető kapcsolattartó
Minden további, az erőforrásokkal és reagensekkel kapcsolatos kérdésben illetékes kapcsolattartó: Qiang Pan-Hammarström (qiang.pan-hammarstrom@ki.se).
Anyagok elérhetősége
Az RBD- és az S1-S2-fehérjék előállíthatók és együttműködés esetén megoszthatók.
Adat- és kódelérhetőség
Az összes releváns adat a cikkben és annak kiegészítő információiban található.
A kísérleti modell és alanyok részletei
A vizsgálat elrendezése és a vizsgálatban résztvevő személyek
A COVID19-donorok szűrését és a mintagyűjtést a Fondazione IRCCS Policlinico San Matteo kórházban, Paviaban, Olaszországban, a COVID19 számára kijelölt egészségügyi intézményben végezték. A vizsgálatban részt vehettek azok, akik elmúltak 18 évesek és hajlandóak és képesek voltak aláírni a betegbeleegyező nyilatkozatot, az E-és RdRp-géneket célzó Corman és mtsai protokolljai alapján valós idejű RT-PCR-rel megerősített SARS-CoV-2-pozitívak voltak és két további, negatív eredményű mintáig történt a megfigyelésük.46 2020. február 28. és október 10. között 78 COVID19-beteget vontak be. Negyvenhét donornál a vérvétel egyetlen időpontban történt, a tünetek kezdete után 7–240 nappal, míg 28 és 3 donornál két, illetve három időpontban vettek vért. A betegség súlyosságának mértéke enyhe (nem kórházi), középsúlyos (hospitalizált, alsó légúti fertőzéssel, nehézlégzéssel vagy anélkül, de oxigéntámogatás nélkül), súlyos (fertőző betegség/szubintenzív osztályon oxigénigénnyel és/vagy pozitív mellkasi komputertomográfia, súlyos alsó légúti fertőzések, bármilyen oxigéntámogatással) és kritikus (intenzív osztályon (ITO) intubált vagy extrakorporális membránoxigenizációs eljárásokkal támogatott betegek) volt.
A betegek demográfiai és klinikai adatait az 1. táblázat tartalmazza és az S1 táblázat részletezi. A betegek medián életkora 63 év volt (32–89 tartomány), 45 (58%) férfi és 33 (42%) nő. A COVID19-klinikai súlyosságának mértéke a kohorszban enyhe (n = 5), középsúlyos (n = 16), súlyos (n = 52) és kritikus (n = 5). A leggyakoribb alapbetegség a magas vérnyomás (40/78, 51%), a cukorbetegség (16/78, 21%), a szívbetegségek (12/78, 15%) és az elhízás (10/78, 13%) volt. A vizsgálat a Policlinico San Matteo kórház intézményi felülvizsgálati testületének jóváhagyásával történt (protokollszám: P_20200029440).
Tizenkét mintát vettek 10 svéd önkéntestől (medián életkor 54 év, tartomány: 29–75), akiknek pozitív volt a PCR-je vagy szeropozitívak voltak SARS-CoV-2-re és enyhe tüneteket tapasztaltak (1. és S1 táblázat). A vérminták gyűjtése a tünetek kezdete után 60–238 nappal történt. A vizsgálatot a stockholmi etikai bizottság hagyta jóvá (Dnr 2020-02646).
Ezenkívül 108 anonimizált (16 és 80 év közötti betegtől származó) szérummintát, amelyet a SARS-CoV-2-pandémia (1995–2005) előtt gyűjtöttek historikus negatív kontrollként használtak fel az ELISA-hoz; és négy egészséges kontrolltól (medián életkor 41 év, 39–50 közötti tartomány) származó PBMC-t és további hét, Svédországban a SARS-CoV-2-pandémia (2011–2020. január) előtt gyűjtött felülúszót használtak fel negatív kontrollként a B- és T-sejt-vizsgálatokhoz. A különböző eljárásokkal vizsgált betegeket és mintákat egy folyamatábrán foglaltuk össze (1. ábra).
Módszerek részletei
SARS-CoV-2-re specifikus antitestek kimutatása
Az RBD-His fehérjét Expi293 sejtekben expresszáltuk és Ni-NTA gyantán (#88221, Thermo Fisher), majd méretkizárásos kromatográfiával Superdex 200 gélfiltrációs oszlopon, PBS-ben tisztítottuk.47 Az S1-S2-His (S-nek nevezett) fehérjét bakulovírus-mentesen High Five rovarsejtekben expresszáltuk48, és a HisTrap excel oszlopon (Cytiva) tisztítottuk, majd preparatív méretkizárásos kromatográfiát végeztünk 16/600 Superdex 200 pg oszlopon (Cytiva).49
A High-binding Corning Half area mikrotiter lemezeket (Corning #3690) egy éjszakán át 4°C-on S- vagy RBD-fehérjével (1,7 μg/ml IgM és IgG esetében; 2,0 μg/ml IgA esetében) PBS-sel inkubáltuk; háromszor mostuk PBS-Tweennel (0,05%), és 2%-os BSA-val blokkoltuk PBS-ben 1 órán át szobahőmérsékleten. A szérumot vagy plazmát 1:3200 (S IgM), 1:6400 (S IgG), 1:1600 (S IgA; RBD IgM, IgA, IgG) mértékben hígítottuk 0,1%-os BSA-ban, majd PBS-ben 1,5 órán át szobahőmérsékleten inkubáltuk. A mikrotiter lemezeket ezután 1 órán át szobahőmérsékleten inkubáltuk torma-peroxidázzal (HRP) konjugált kecske anti-humán IgM-mel (Invitrogen #A18835), kecske anti-humán IgA-val (Jackson #109-036-011) vagy kecske anti-humán IgG-vel (Invitrogen #A18805) (mindegyik 1:15 000 arányban 0,1%-os BSA-PBS-ben hígítva). A megkötött antitesteket tetrametilbenzidin szubsztrát (Sigma #T0440) segítségével detektáltuk. A színreakciót 0,5M H2SO4 használatával állítottuk le. Az abszorbanciát 450 nm-en mértük. Az antitestszinteket tetszőleges egységekben (AU/ml) tüntettük fel, a nagy mértékben pozitív szérumkészlet sorozathígítással készített standard görbéje alapján. Az antitest-pozitivitás küszöbértékét minden antigénre és izotípusra meghatároztuk hatásfokmérő karakterisztika (ROC) görbék használatával a historikus kontrollokban (n = 108) és a COVID19-betegek antitestválaszai alapján (55 mintát gyűjtöttünk 7–28 nappal a tünetek kezdete után). A beállított pozitivitási küszöbérték anti-S IgM esetén >2,5 AU/ml, anti-S IgA esetén >0,5 AU/ml, anti-S IgG esetén >0,03 AU/ml, anti-RBD IgM esetén >8,4 AU/ml, anti-RBD IgA esetén > 0,08 AU/ml és anti-RBD IgG esetén > 14,8 AU/ml, 96%-os IgM-, 99%-os IgA- és 97%-os IgG-specifitást adva. Egy korábban leírt mikroneutralizációs eljárást50,51 alkalmaztunk a SARS-CoV-2-NT-antitesttiterek meghatározására 96 mintában. A neutralizációs titer az a maximális hígítás volt, amely a citopátiás hatás 90%-os csökkenését eredményezte.
PBMC- és RNS-izolálás
A PBMC-ket a vérből vagy felülúszóból izoláltuk standard sűrűséggradiens centrifugálással Lymphoprep (Axis-Shield) alkalmazásával, és mélyfagyasztott állapotban folyékony nitrogénben tároltuk az elemzésig. A teljes RNS-t PBMC-ből RNeasy mini kit alkalmazásával izoláltuk a gyártó protokollja (QIAGEN) szerint.
A SARS-CoV-2-elleni B- és T-sejt-válasz
A PBMC-k négy napig 10%-os FCS-t tartalmazó RPMI-1640 médiumban inkubáltuk TLR7- és TLR8-agonista imidazoquinolin rezikimoddal (R848, 1 μg/ml; Mabtech AB, Nacka, Svédország) és rekombináns humán IL-2-vel (10 ng/ml) kiegészítve a memória B-sejtek stimulálására.52 A befogó monoklonális anti-humán IgG-antitestekkel előzetesen bevont ELISpot mikrotiter lemezeket összesen 300 000 illetve 30 000 előzetesen stimulált sejttel inkubáltuk lyukanként RBD-specifikus IgG-t, illetve az össz IgG-t szekretáló sejt detektálására. A SARS-CoV-2 RBD-specifikus IgG-antitesteket szekretáló B-sejtek és az IgG-t (össz IgG) szekretáló sejtek számát humán IgG SARS-CoV-2 RBD ELISpotPLUS (ALP) kit alkalmazásával mértük a gyártó protokollja szerint (Mabtech AB).
Az IFN-γ-t és IL-2-t szekretáló T-sejteket humán IFN-γ/IL-2 SARS-CoV-2 FluoroSpotPLUS kit használatával detektáltuk a gyártó protokollja szerint (Mabtech AB). A befogó monoklonális anti-IFN-y-val és anti-IL-2-vel előzetesen bevont mikrotiter lemezeket egy éjszakán át RPMI-1640 médiumban inkubáltuk, amely 10%-os FCS-t tartalmazott, illetve az alábbi keverékkel egészítettük ki: SARS-CoV-2-peptidkészlet (pásztázó vagy meghatározott készletek), anti-CD28 (100 ng/ml) és lyukanként 300 000 sejt párásított inkubátorokban (5% CO2, 37°C).
A SARS-CoV-2 S1-pásztázó készlet 166 peptidet tartalmaz a humán SARS-CoV-2-vírusból (#3629-1, Mabtech AB). A peptidek 15-merek, amelyek 11 aminosavas átfedésben vannak, lefedik az S-fehérje S1-doménjét (13–685 aminosav). A SARS-CoV-2 S N M O által meghatározott peptidkészlet 47 szintetikus peptidet tartalmaz, amely az S, N, M ORF3a és ORF7a fehérjékből származó humán HLA-hoz kötődik (#3622-1, Mabtech AB).45 A SARS-CoV-2 S2 N által definiált peptidkészlet 41 szintetikus peptidet tartalmaz, amely a SARS-CoV-2-vírus S- és N-fehérjékből származó humán HLA-hoz kötődik (#3620-1, Mabtech AB).53 Az ELISpot és a Fluorospot eljárások eredményeit IRIS-reader segítségével értékeltük, és az IRIS 1.1.9-es verziójával elemeztük (Mabtech AB). Az eredményeket a negatív kontroll hátterének levonása után 300 000 szélesztett sejtre jutó pöttyök számaként fejeztük ki. A küszöbértéket a negatív kontrollokból a specifikus B- és T-sejtek legmagasabb mennyiségére állítottuk be.
Kvantifikálás és statisztikai elemzés
Mann–Whitney U-próbával történt a SARS-CoV-2-ellenes antitestszintek és a specifikus memória B- és T-sejtek számának összehasonlítása a csoportok között. A korrelációs elemzést Spearman-rangkorrelációval végeztük. Az illesztett minták összehasonlításához Wilconxon-féle előjeles rangpróbát használtunk. Minden elemzést és adatábrázolást GraphPad vagy 3.6.1 verziószámú R szoftverrel készítettünk.
Köszönetnyilvánítás
Köszönjük a betegek, a donorok és a klinikusok hozzájárulását. A projekt létrejöttét az Európai Unió Horizont 2020 kutatási és innovációs keretprogramja (ATAC, 101003650), az Italian Ministry of Health (olasz egészségügyi minisztérium, Ricerca Finalizzata, támogatás száma: GR-2013-02358399), a Center for Innovative Medicine (innovatív orvosi központ), és a Swedish Research Council (svéd kutatási tanács) támogatta. J.A. a SciLifeLab/KAW a 2020-as nemzeti COVID19 kutatási program projektjének támogatásában részesült.
Szerzői hozzájárulások
N.S. végezte el az ELISA-kísérletet. A.P., I.C., E.P., A.S., F. Bergami, R.D., H.A., L.D., M. Sachs és F. Baldanti segített a betegadatok gyűjtésében, a minta előkészítésében és a neutralizációs vizsgálatban. L.D., M.K.-B. és S.B.-A. tervezte meg és/vagy végezte el az ELISpot- és a FluoroSpot-kísérleteket. M.V., M. Sambo, V.Z., R.B., T.O. és F.M. részt vett a betegek bevonásában, a betegek kezelésében és a klinikai adatok gyűjtésében. J. A., F. Bertoglio, M. Schubert és M. H. végezte a fehérjék előállítását és tisztítását. N.S., L.D., H.W., M.K.-B., M.B.-S., J.H., H.A., L.H., H.M. és Q.P.-H. segített az adatok elemzésében és értelmezésében. N.S., H.M., és Q.P.-H. készítette el a kéziratot. Y.X., L.H., H.M., F.B., és Q.P.-H. tervezte meg és felügyelte a vizsgálatot.
Összeférhetetlenségi nyilatkozatok
S.B.-A. tagja a Mabtech tanácsadó testületének. A többi szerzőnél nem áll fenn összeférhetetlenség.
Kiegészítő információk
Irodalomjegyzék
- Naqvi A.A.T. Fatima K. Mohammad T. Fatima U. Singh I.K. Singh A. tif S.M. Hariprasad G. Hasan G.M. Hassan M.I.
Insights into SARS-CoV-2 genome, structure, evolution, pathogenesis and therapies: structural genomics approach.Biochim. Biophys. Acta Mol. Basis Dis. 2020; 1866: 165878 - Grifoni A. Weiskopf D. Ramirez S.I. Mateus J. Dan J.M. Moderbacher C.R. Rawlings S.A. Sutherland A. Premkumar L. Jadi R.S.et al.
Targets of T cell responses to SARS-CoV-2 coronavirus in humans with COVID-19 disease and unexposed individuals.Cell. 2020; 181: 1489-1501.e15 - Juno J.A. Tan H.-X. Lee W.S. Reynaldi A. Kelly H.G. Wragg K. Esterbauer R. Kent H.E. Batten C.J. Mordant F.L. et al.
Humoral and circulating follicular helper T cell responses in recovered patients with COVID-19.Nat. Med. 2020; 26: 1428-1434 - Robbiani D.F. Gaebler C. Muecksch F. Lorenzi J.C.C. Wang Z. Cho A. Agudelo M. Barnes C.O. Gazumyan A. Finkin S.et al.
Convergent antibody responses to SARS-CoV-2 in convalescent individuals.Nature. 2020; 584: 437-442 - Hassan A.O. Case J.B. Winkler E.S. Thackray L.B. Kafai N.M. Bailey A.L. McCune B.T. Fox J.M. Chen R.E. Alsoussi W.B.et al.
A SARS-CoV-2 infection model in mice demonstrates protection by neutralizing antibodies.Cell. 2020; 182: 744-753.e4 - Rogers T.F. Zhao F. Huang D. Beutler N. Burns A. He W.-T. Limbo O. Smith C. Song G. Woehl J. et al.Iso
lation of potent SARS-CoV-2 neutralizing antibodies and protection from disease in a small animal model.Science. 2020; 369: 956-963 - DeFrancesco L.
COVID-19 antibodies on trial.Nat. Biotechnol. 2020; 38: 1242-1252 - Chen P. Nirula A. Heller B. Gottlieb R.L. Boscia J. Morris J. Huhn G. Cardona J. Mocherla B. Stosor V. et al.
BLAZE-1 InvestigatorsSARS-CoV-2 Neutralizing Antibody LY-CoV555 in Outpatients with Covid-19.N. Engl. J. Med. 2021; 384: 229-237 - Hammarström L. Abolhassani H. Baldanti F. Marcotte H. Pan-Hammarström Q.
Development of passive immunity against SARS-CoV-2 for management of immunodeficient patients-a perspective.J. Allergy Clin. Immunol. 2020; 146: 58-60 - Chandrashekar A. Liu J. Martinot A.J. McMahan K. Mercado N.B. Peter L. Tostanoski L.H. Yu J. Maliga Z. Nekorchuk M. et al.
SARS-CoV-2 infection protects against rechallenge in rhesus macaques.Science. 2020; 369: 812-817 - Gudbjartsson D.F. Norddahl G.L. Melsted P. Gunnarsdottir K. Holm H. Eythorsson E. Arnthorsson A.O. Helgason D. Bjarnadottir K. Ingvarsson R.F. et al.
Humoral immune response to SARS-CoV-2 in Iceland.N. Engl. J. Med. 2020; 383: 1724-1734 - Ibarrondo F.J. Fulcher J.A. Goodman-Meza D. Elliott J. Hofmann C. Hausner M.A. Ferbas K.G. Tobin N.H. Aldrovandi G.M. Yang O.O.
Rapid decay of anti-SARS-CoV-2 antibodies in persons with mild Covid-19.N. Engl. J. Med. 2020; 383: 1085-1087 - Long Q.-X. Tang X.-J. Shi Q.-L. Li Q. Deng H.-J. Yuan J. Hu J.-L. Xu W. Zhang Y. Lv F.-J. et al.
Clinical and immunological assessment of asymptomatic SARS-CoV-2 infections.Nat. Med. 2020; 26: 1200-1204 - Bölke E. Matuschek C. Fischer J.C.
Loss of anti-SARS-CoV-2 antibodies in mild covid-19.N. Engl. J. Med. 2020; 383: 1694-1695 - Iyer A.S. Jones F.K. Nodoushani A. Kelly M. Becker M. Slater D. Mills R. Teng E. Kamruzzaman M. Garcia-Beltran W.F. et al.
Persistence and decay of human antibody responses to the receptor binding domain of SARS-CoV-2 spike protein in COVID-19 patients.Sci. Immunol. 2020; 5: eabe0367 - Isho B. Abe K.T. Zuo M. Jamal A.J. Rathod B. Wang J.H. Li Z. Chao G. Rojas O.L. Bang Y.M. et al.
Persistence of serum and saliva antibody responses to SARS-CoV-2 spike antigens in COVID-19 patients.Sci. Immunol. 2020; 5: eabe55115 - Wajnberg A. Amanat F. Firpo A. Altman D.R. Bailey M.J. Mansour M. McMahon M. Meade P. Mendu D.R. Muellers K. et al.
Robust neutralizing antibodies to SARS-CoV-2 infection persist for months.Science. 2020; 370: 1227-1230 - Hartley G.E. Edwards E.S.J. Aui P.M. Varese N. Stojanovic S. McMahon J. Peleg A.Y. Boo I. Drummer H.E. Hogarth P.M. et al.
Rapid generation of durable B cell memory to SARS-CoV-2 spike and nucleocapsid proteins in COVID-19 and convalescence.Sci. Immunol. 2020; 5: eabf8891 - Röltgen K. Powell A.E. Wirz O.F. Stevens B.A. Hogan C.A. Najeeb J. Hunter M. Wang H. Sahoo M.K. Huang C. et al.
Defining the features and duration of antibody responses to SARS-CoV-2 infection associated with disease severity and outcome.Sci. Immunol. 2020; 5: eabe0240 - Dan J.M. Mateus J. Kato Y. Hastie K.M. Yu E.D. Faliti C.E. Grifoni A. Ramirez S.I. Haupt S. Frazier A. et al.
Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection.Science. 2021; 371: eabf4063 - Kutsuna S. Asai Y. Matsunaga A.
Loss of anti–SARS-CoV-2 antibodies in mild covid-19.N. Engl. J. Med. 2020; 383: 1695-1696 - Huang A.T. Garcia-Carreras B. Hitchings M.D.T. Yang B. Katzelnick L.C. Rattigan S.M. Borgert B.A. Moreno C.A. Solomon B.D. Trimmer-Smith L. et al.
A systematic review of antibody mediated immunity to coronaviruses: kinetics, correlates of protection, and association with severity.Nat. Commun. 2020; 11: 4704 - Zheng S. Fan J. Yu F. Feng B. Lou B. Zou Q. Xie G. Lin S. Wang R. Yang X. et al.
Viral load dynamics and disease severity in patients infected with SARS-CoV-2 in Zhejiang province, China, January-March 2020: retrospective cohort study.BMJ. 2020; 369: m1443 - Bastard P. Rosen L.B. Zhang Q. Michailidis E. Hoffmann H.-H. Zhang Y. Dorgham K. Philippot Q. Rosain J. Béziat V. et al.
- HGID Lab
- NIAID-USUHS Immune Response to COVID Group
- COVID Clinicians
- COVID-STORM Clinicians
- Imagine COVID Group
- French COVID Cohort Study Group
- Milieu Intérieur Consortium
- CoV-Contact Cohort
- Amsterdam UMC Covid-19 Biobank
- COVID Human Genetic EffortAutoantibodies against type I IFNs in patients with life-threatening COVID-19.Science. 2020; 370: eabd4585
- View in Article
- Zhang Q. Bastard P. Liu Z. Le Pen J. Moncada-Velez M. Chen J. Ogishi M. Sabli I.K.D. Hodeib S. Korol C. et al.
- COVID-STORM Clinicians
- COVID Clinicians
- Imagine COVID Group
- French COVID Cohort Study Group
- CoV-Contact Cohort
- Amsterdam UMC Covid-19 Biobank
- COVID Human Genetic Effort
- NIAID-USUHS/TAGC COVID Immunity GroupInborn errors of type I IFN immunity in patients with life-threatening COVID-19.Science. 2020; 370: eabd4570
- View in Article
- Yang O.O. Ibarrondo F.J.
Loss of Anti-SARS-CoV-2 Antibodies in Mild Covid-19. Reply.N. Engl. J. Med. 2020; 383: 1697-1698 - Premkumar L. Segovia-Chumbez B. Jadi R. Martinez D.R. Raut R. Markmann A. Cornaby C. Bartelt L. Weiss S. Park Y. et al.
The receptor binding domain of the viral spike protein is an immunodominant and highly specific target of antibodies in SARS-CoV-2 patients.Sci. Immunol. 2020; 5: eabc8413 - Trinité B. Tarrés-Freixas F. Rodon J. Pradenas E. Urrea V. Marfil S. Rodríguez de la Concepción M.L. Ávila-Nieto C. Aguilar-Gurrieri C.Barajas A. et al.
SARS-CoV-2 infection elicits a rapid neutralizing antibody response that correlates with disease severity.Sci. Rep. 2021; 11: 2608 - Rodda L.B. Netland J. Shehata L. Pruner K.B. Morawski P.A. Thouvenel C.D. Takehara K.K. Eggenberger J. Hemann E.A. Waterman H.R. et al.
Functional SARS-CoV-2-specific immune memory persists after mild COVID-19.Cell. 2021; 184: 169-183.e17 - Vaisman-Mentesh A. Dror Y. Tur-Kaspa R. Markovitch D. Kournos T. Dicker D. Wine Y.
SARS-CoV-2 specific memory B cells frequency in recovered patient remains stable while antibodies decay over time.medRxiv. 2020; https://doi.org/10.1101/2020.08.23.20179796 - Wheatley A.K. Juno J.A. Wang J.J. Selva K.J. Reynaldi A. Tan H.-X. Lee W.S. Wragg K.M. Kelly H.G. Esterbauer R. et al.
Evolution of immunity to SARS-CoV-2.medRxiv. 2020; https://doi.org/10.1101/2020.09.09.20191205 - Gaebler C. Wang Z. Lorenzi J.C.C. Muecksch F. Finkin S. Tokuyama M. Cho A. Jankovic M. Schaefer-Babajew D. Oliveira T.Y. et al.
Evolution of antibody immunity to SARS-CoV-2.Nature. 2021; https://doi.org/10.1038/s41586-021-03207-w - Zhang F. Gan R. Zhen Z. Hu X. Li X. Zhou F. Liu Y. Chen C. Xie S. Zhang B. et al.
Adaptive immune responses to SARS-CoV-2 infection in severe versus mild individuals.Signal Transduct. Target. Ther. 2020; 5: 156 - Sekine T. Perez-Potti A. Rivera-Ballesteros O. Strålin K. Gorin J.-B. Olsson A. Llewellyn-Lacey S. Kamal H. Bogdanovic G. Muschiol S. et al.
Karolinska COVID-19 Study GroupRobust T cell immunity in convalescent individuals with asymptomatic or mild COVID-19.Cell. 2020; 183: 158-168.e14 - Braun J. Loyal L. Frentsch M. Wendisch D. Georg P. Kurth F. Hippenstiel S. Dingeldey M. Kruse B. Fauchere F. et al.
SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19.Nature. 2020; 587: 270-274 - Zuo J. Dowell A. Pearce H. Verma K. Long H. Begum J. Aiano F. Amin-Chowdhury Z. Hallis B. Stapley L. et al.
Robust SARS-CoV-2-specific T-cell immunity is maintained at 6 months following primary infection.bioRxiv. 2020; https://doi.org/10.1101/2020.11.01.362319 - Breton G. Mendoza P. Hägglöf T. Oliveira T.Y. Schaefer-Babajew D. Gaebler C. Turroja M. Hurley A. Caskey M. Nussenzweig M.C.Persistent cellular immunity to SARS-CoV-2 infection.J. Exp. Med. 2020; 218: e20202515
- Bert N.L. Clapham H.E. Tan A.T. hia W.N. Tham C.Y. Lim J.M. Kunasegaran K. Tan L. Dutertre C.-A. Shankar N. et al.
Highly functional virus-specific cellular immune response in asymptomatic SARS-CoV-2 infection.bioRxiv. 2020; https://doi.org/10.1101/2020.11.25.399139 - Nelde A. Bilich T. Heitmann J.S. Maringer Y. Salih H.R. Roerden M. Lübke M. Bauer J. Rieth J. Wacker M. et al.
SARS-CoV-2-derived peptides define heterologous and COVID-19-induced T cell recognition.Nat. Immunol. 2021; 22: 74-85 - Schwarzkopf S. Krawczyk A. Knop D. Klump H. Heinold A. Heinemann F.M. Thümmler L. Temme C. Breyer M. Witzke O. et al.
Cellular Immunity in COVID-19 convalescents with PCR-confirmed infection but with undetectable SARS-CoV-2–specific IgG.Emerg. Infect. Dis. 2021; 27 https://doi.org/10.3201/eid2701.203772 - Yu J. Tostanoski L.H. Peter L. Mercado N.B. McMahan K. Mahrokhian S.H. Nkolola J.P. Liu J. Li Z. Chandrashekar A. et al.
DNA vaccine protection against SARS-CoV-2 in rhesus macaques.Science. 2020; 369: 806-811 - Edridge A.W.D. Kaczorowska J. Hoste A.C.R. Bakker M. Klein M. Loens K. Jebbink M.F. Matser A. Kinsella C.M. Rueda P. et al.
Seasonal coronavirus protective immunity is short-lasting.Nat. Med. 2020; 26: 1691-1693 - Callow K.A. Parry H.F. Sergeant M. Tyrrell D.A.
The time course of the immune response to experimental coronavirus infection of man.Epidemiol. Infect. 1990; 105: 435-446 - Callaway E.
Fast-spreading COVID variant can elude immune responses.Nature. 2021; 589: 500-501 - Peng Y. Mentzer A.J. Liu G. Yao X. Yin Z. Dong D. Dejnirattisai W. Rostron T. Supasa P. Liu C. et al.
Oxford Immunology Network Covid-19 Response T cell Consortium
ISARIC4C InvestigatorsBroad and strong memory CD4+ and CD8+ T cells induced by SARS-CoV-2 in UK convalescent individuals following COVID-19.Nat. Immunol. 2020; 21: 1336-1345 - Corman V.M. Landt O. Kaiser M. Molenkamp R. Meijer A. Chu D.K. Bleicker T. Brünink S. Schneider J. Schmidt M.L. et al.
Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR.Euro Surveill. 2020; 25: 2000045 - Amanat F. Stadlbauer D. Strohmeier S. Nguyen T.H.O. Chromikova V. McMahon M. Jiang K. Arunkumar G.A. Jurczyszak D. Polanco J. et al.
A serological assay to detect SARS-CoV-2 seroconversion in humans.Nat. Med. 2020; 26: 1033-1036 - Bleckmann M. Schürig M. Endres M. Samuels A. Gebauer D. Konisch N. van den Heuvel J.
Identifying parameters to improve the reproducibility of transient gene expression in High Five cells.PLoS ONE. 2019; 14: e0217878 - Bertoglio F. Meier D. Langreder N. Steinke S. Rand U. Simonelli L. Heine P.A. Ballmann R. Schneider K.-T. Roth K.D.R. et al.
SARS-CoV-2 neutralizing human recombinant antibodies selected from pre-pandemic healthy donors binding at RBD-ACE2 interface.bioRxiv. 2020; https://doi.org/10.1101/2020.06.05.135921 - Percivalle E. Cambiè G. Cassaniti I. Nepita E.V. Maserati R. Ferrari A. Di Martino R. Isernia P. Mojoli F. Bruno R. et al.
Prevalence of SARS-CoV-2 specific neutralising antibodies in blood donors from the Lodi Red Zone in Lombardy, Italy, as at 06 April 2020.Euro Surveill. 2020; 25: 2001031 - Bonelli F. Sarasini A. Zierold C. Calleri M. Bonetti A. Vismara C. Blocki F.A. Pallavicini L. Chinali A. Campisi D. et al.
Clinical and analytical performance of an automated serological test that identifies S1/S2-neutralizing IgG in COVID-19 patients semiquantitatively.J. Clin. Microbiol. 2020; 58 (e01224-20) - Jahnmatz M. Kesa G. Netterlid E. Buisman A.-M. Thorstensson R. Ahlborg N.
Optimization of a human IgG B-cell ELISpot assay for the analysis of vaccine-induced B-cell responses.J. Immunol. Methods. 2013; 391: 50-59 - Ahmed S.F. Quadeer A.A. McKay M.R.
Preliminary identification of potential vaccine targets for the COVID-19 coronavirus (SARS-CoV-2) based on SARS-CoV immunological studies.Viruses. 2020; 12: 254
Információk a cikkről
A publikáció története
Közzétéve: 2021. február 10.
Elfogadva: 2021. február 3.
Érkezett felülvizsgált formában: 2020. december 28.
Érkezett: 2020. november 21.
Azonosítás
DOI: https://doi.org/10.1016/j.medj.2021.02.001
Szerzői jog
© 2021 A Szerző(k). Közzétette: Elsevier Inc.
Felhasználói licenc
Creative Commons Attribution (CC BY 4.0) |
Hogyan használható fel újra
ScienceDirect
Hozzáférés a cikkhez: ScienceDirect
{kind=link}